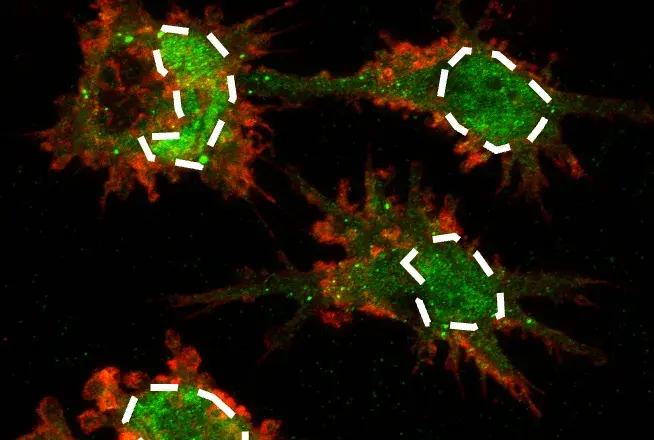
Mitochondrial health is a complex, carefully regulated process involving pathways that determine mitochondria size, biogenesis, and degradation. Mitochondrial size is a dynamic process involving both fusion (enlargement) and fission (shrinkage). Elimination of dysfunctional mitochondria, typically associated with fission, occurs through a selective pathway of autophagy called mitophagy. Autophagy is a catabolic process for the clearance of cytoplasmic contents captured by the autophagosome and degraded by lysosomes. While autophagy may be nonselective, it can also be used to specifically target various organelles, pathogens, and even proteins.
Mitophagy is the best-studied example of selective autophagy involving several mitochondrial-localized cargo receptors accommodating cues such as mitochondrial depolarization, oxidative stress, and hypoxia. Cargo receptors containing an LC3-interacting region (LIR) interact with LC3 family members, bridging targets to autophagosomes and subsequent degradation by the lysosome.
The PINK1/Parkin Pathway
The best-studied mitophagy pathway involves PINK1/Parkin activation following mitochondrial depolarization. PINK1 is a mitochondrial serine/threonine kinase that is normally kept at low levels upon translocation through the inner mitochondrial membrane, where it is cleaved by the peptidase PARL (presenilin-associated rhomboid-like protein) and targeted for proteasomal degradation. Inhibition of mitochondrial membrane potential, preventing translocation of PINK1, results in stabilization on the outer mitochondrial membrane, phosphorylation of Ser65 of ubiquitin, and recruitment and phosphorylation of the E3 ligase parkin.
Activation of Parkin leads to the accumulation of ubiquitinated chains on multiple mitochondrial proteins that signal ubiquitin-binding cargo receptors, including SQSTM1/p62, Optineurin, and NDP52. Also important in this process is the phosphorylation of cargo receptors by TBK1, a kinase involved in innate immunity that is recruited to the mitochondria and enhances mitophagy.
Blog: It’s a Cell-Eat-Self World: ULK1, AMPK & mTOR Signaling in Autophagy
Independent of PINK1, several mitochondrial cargo receptors have been described, including BNIP3, BNIP3L/Nix, FUNDC1, BCL2L13, and FKBP8. Hypoxia-induced mitophagy involves BNIP3, BNIP3L/Nix, and FUNDC1. Both BNIP3 and BNIP3L/Nix are transcriptionally induced by HIF1α and contain LIR domains for recruitment to the autophagosome. BNIP3L/Nix is also important during reticulocyte development, in which mitochondria are depleted. FUNDC1, another cargo receptor important for hypoxia-induced mitophagy, is regulated by the autophagy kinase ULK1. The interplay of these and other cargo receptors is of growing interest.
Implications in Disease and Neurodegeneration
Excessive or inadequate mitophagy has been described as a contributing factor to many pathological conditions, including neurodegeneration, metabolic disorders, muscular dystrophy, liver disease, cardiovascular disease, and cancer. Defects in mitophagy lead to an accumulation of damaged mitochondria, which leads to an accumulation of reactive oxygen species (ROS) and cell death. Perhaps the clearest example was illustrated with the genetic mutations of PINK1 and PARK2 (encoding Parkin) found in early-onset Parkinson’s disease. These genetic events implicate the loss of mitophagy and accumulation of dysfunctional mitochondria in this disease.
Blog: Unraveling Parkinson’s Disease: The Role of Mitophagy, Autophagy & Lysosomal Processing
Similarly, mutations in OPTN (encoding Optineurin), SQSTM1, and TBK1 have been identified in other neurodegenerative diseases including Amyotrophic Lateral Sclerosis (ALS). Accumulation of dysfunctional mitochondria due to inadequate mitophagy is also found in other diseases associated with cell death, though the direct roles of mitophagy in many cases are not clear.
Assay Tools to Study Mitophagy and Advance Biomarker Discovery
To help further elucidate the role of mitophagy in health and disease, a number of tools have been developed to study this process. Mt-Keima and Mito-QC are pH-sensitive mitochondrial fluorescent probes to monitor mitophagy and have been utilized with in vivo models, but have limited utility to study pathological samples. It is also possible to use immunofluorescent staining to monitor the co-localization of proteins expressed on the mitochondria and autophagosomes, but these studies tend not to be quantitative.
The discovery of regulated mitophagy pathways is paving the way for valuable biomarker discovery and ways to monitor mitophagy. Specific antibodies can be used to monitor changes in phosphorylated Optineurin as well as changes in PINK1 and its substrates like ubiquitin. ELISA assays developed to detect Phospho-Ubiquitin (Ser65) have been shown to be sensitive and specific and able to measure changes in mitophagy found in neurodegeneration including Parkinson’s and Alzheimer’s diseases. These assays may even prove sensitive enough to observe mitophagy differences in plasma samples.
Cell Signaling Technology has developed validated antibodies and kits designed to meet the needs of mitophagy researchers and progress therapeutic development.