If the thought of self-cannibalization is not appealing to you, you may not want to read the next sentence... Your cells are literally eating themselves right now! In the 1960s, Nobel Prize-winning Belgian cytologist and biochemist Christian de Duve named this process autophagy from the Greek auto (self) and phagein (to eat). Little did de Duve know what an issue this word would cause: Do you pronounce it “aw-tof-a-gee” or, “auto-fay-gee”? Well, actually—it’s a bit like "tomayto, tomahto"—either pronunciation is acceptable. As de Duve himself said, “I coined the word, but not the pronunciation.”
In this post, we explore the mechanisms of autophagy and the role of the ULK1 complex, AMPK, and mTOR, in stress-induced autophagy signaling.
Autophagy Signaling Pathway and Mechanisms
However you say it, autophagy allows bulk cytoplasmic contents, abnormal proteins, and old or damaged organelles to be broken down and the resulting metabolites to be recycled. It occurs at a basal rate in “normal” cells to maintain homeostasis, but it is also a survival mechanism employed during stressful conditions like nutrient starvation, oxygen depletion, and endoplasmic reticulum (ER) stress. During conditions such as these, autophagy is induced to provide cells with much-needed nutrients.1 In simplified terms, you could say that autophagy is a cell's way of "reducing, reusing, and recycling" when resources become scarce—with emphasis on the reusing and recycling part of that phrase!
Blog: Characterizing the Autophagic-Lysosomal Pathway in Parkinson’s Disease
In response to stress, such as nutrient starvation, i.e., amino acid or glucose depletion, a double membrane structure called a phagophore forms within the cell, which elongates and starts to encompass the cytoplasmic components (cargo). Eventually, the membrane seals around the cargo, forming a structure called the autophagosome. The autophagosome then fuses with a lysosome, a membrane-bound organelle that contains a mixture of enzymes that work best in acidic conditions, to form an autolysosome or autophagolysosome. The cargo undergoes degradation by lysosomal enzymes and the resulting nutrients become available for the cell to reuse.

Autophagolysosome formation. View the interactive Autophagy Signaling Pathway on the CST website.
Historically, a lot of what we have learned about mammalian autophagy signaling came from studying yeast.2 And, as it turns out, there is a high degree of conservation in autophagy mechanisms between yeast and mammals. Fifteen autophagy-related (Atg) genes were first identified in a genetic screen performed on yeast, and mutations in these genes caused defects in autophagy.3 Now, at least 41 Atg genes have been reported in yeast.4 Many of the autophagy-related proteins in mammals are named for their yeast counterparts, which is why a lot of proteins in the mammalian autophagy pathway bear the name “AtgX”.
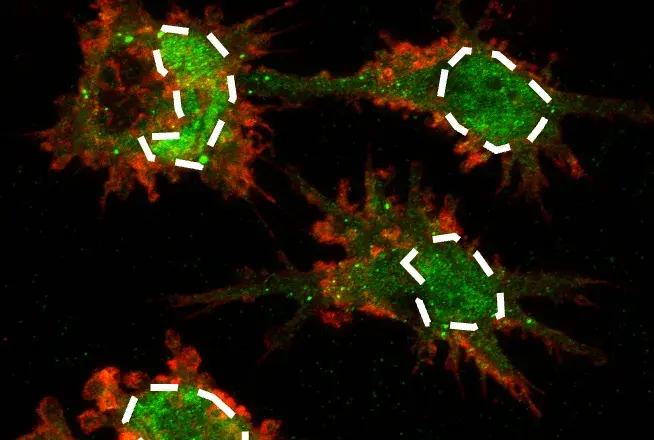
.png?width=600&height=300&name=Autophagy_mTOR%20inhibitor_Phospho-Atg14%20(Ser29).png) Immunofluorescence (IF) analysis of HCT 116 Atg14 cells untreated (left) and treated with mTOR-inhibitor Torin 1 #14379 (right), using Phospho-Atg14 (Ser29) (D4B8M) Rabbit mAb #92340 (green). In this experiment, the low level of expression of phosphorylated Atg14 in the left image suggests that autophagy is "off". After treatment with Torin 1, Atg14 is phosphorylated, mTOR activity is inhibited, and autophagy is "on", as depicted in the right image. For more data images detailing the validation experiments for #92340, including knockout validation testing in IF, visit the product page for #92340.
Immunofluorescence (IF) analysis of HCT 116 Atg14 cells untreated (left) and treated with mTOR-inhibitor Torin 1 #14379 (right), using Phospho-Atg14 (Ser29) (D4B8M) Rabbit mAb #92340 (green). In this experiment, the low level of expression of phosphorylated Atg14 in the left image suggests that autophagy is "off". After treatment with Torin 1, Atg14 is phosphorylated, mTOR activity is inhibited, and autophagy is "on", as depicted in the right image. For more data images detailing the validation experiments for #92340, including knockout validation testing in IF, visit the product page for #92340.
We now know that autophagy is involved in many physiological and pathological events including cardiovascular, neurodegenerative, metabolic, musculoskeletal, pulmonary, and infectious diseases, and we are making great strides toward understanding the complex, highly regulated signaling involved in autophagy.
Autophagy Regulation: The Role of the ULK1 Complex
Prior to autophagosome assembly, the autophagy signaling pathway is initiated by activation of the ULK1 complex in mammals, or the ATG1 complex in yeast, which is comprised of ULK1 or ULK2, FIP200, Atg101, and Atg13.5 The ULK1 complex acts as a bridge between the nutrient and energy sensors mTOR (mammalian target of rapamycin) and AMPK upstream, and the formation of autophagosomes downstream.
Related: AMPK Signaling Pathway and mTOR Signaling Pathway
ULK1 and ULK2 are highly regulated by multiple phosphorylation sites.6 When activated, the ULK1 complex associates with phagophores to form ULK1 puncta on the phagophore membrane, where several other complexes are then recruited in a process that is still poorly understood.7
ULK1 Phosphorylation by AMPK and mTOR
It has long been known that the phosphorylation of ULK1 is a key mechanism for autophagy regulation. One widely accepted theory is that nutrient-starved cells can induce autophagy through AMPK, while mTOR prevents autophagy under nutrient-rich conditions.

IF analysis of HeLa cells treated with Rapamycin #9904, an mTOR-inhibitor (left) or treated with insulin, which activates mTOR (right), using Phospho-mTOR (Ser2448) (D9C2) XP® Rabbit mAb #5536 (green). The lack of phospho-mTOR in the left image suggests that autophagy is "on". After rapamycin treatment, the right image shows that mTOR has been phosphorylated at Ser2448, which suggests that autophagy has been downregulated (i.e., turned "off").
When AMPK phosphorylates ULK1 at Ser 317, Ser 467, Ser 555, Ser 574, Ser 638, and Ser 777, the ULK complex is activated, promoting or turning "on" autophagy. Conversely, when nutrients are plentiful, AMPK is inactive, and mTOR associates with and phosphorylates ULK1 at Ser 757, preventing ULK1 activation by AMPK. In this situation, autophagy is “turned off”.8,9

IF analysis of MCF7 cells untreated (left); or treated with mTOR-inhibitor Torin 1 #14379 (right) using Phospho-ULK1 (Ser757) (D7O6U) Rabbit mAb #14202 (green). This experiment suggests that autophagy is "off" in the left image, which depicts cells from the breast cancer cell line MCF7. After mTOR activity is inhibited, autophagy is turned back "on", as depicted in the right image.
However, recent studies continue to uncover additional hypotheses that offer alternative methods of regulating autophagy. In a 2023 research paper by JM Park et al., AMPK was found to inhibit ULK complex activity by phosphorylating ULK1 at Thr 660.10 This finding suggests a potential dual function of AMPK that would change our understanding of autophagy signaling and cellular homeostasis during stress.
As well as being at the hub of autophagy signaling, the ULK complex is notable for being the only serine/threonine kinase among the core autophagy proteins. In 2015, Egan et al. reported a ULK1 consensus phosphorylation motif.11 To identify ULK1 substrates, they searched the human proteome for proteins containing this motif. Not surprisingly, many core autophagy proteins were found.
The following ULK1 substrates represent the most reliable phosphorylation sites currently known to regulate autophagy:
Unraveling the autophagy signaling pathway will help us to more clearly understand the role of autophagy in maintaining cell homeostasis and how it is misregulated in diseases like cancer, diabetes, and neurodegeneration.
Could inhibiting autophagy by inactivating essential proteins like ULK1 be a therapeutic possibility for these diseases? It definitely provides some food for thought.
Autophagy Signaling Antibody Sampler Kits
Check out our antibody sampler kits, which contain the reagents you need to drive your research forward:
- Autophagy Induction (ULK1 Complex) Antibody Sampler Kit #46486
- ULK1 Substrate Antibody Sampler Kit #85493
- Autophagy Antibody Sampler Kit #4445
- Human Reactive Cell Death and Autophagy Antibody Sampler Kit #42867
- Mouse Reactive Cell Death and Autophagy Antibody Sampler Kit #48734
- ER Stress-induced Autophagy Antibody Sampler Kit #89947
- Autophagy Vesicle Nucleation Antibody Sampler Kit #70751
- Autophagy Atg8 Family Antibody Sampler Kit #64459
- Autophagy Vesicle Elongation (LC3 Conjugation) Antibody Sampler Kit #19848
- Autophagy Vesicle Elongation (Atg12 Conjugation) Antibody Sampler Kit #32345
Additional Resources
Interested in learning more about signaling in autophagy? Access our comprehensive list of Cell Growth & Viability Signaling Pathways, which includes the following pathways: Autophagy Signaling, PI3K/Akt Signaling, p53 Signaling, MAPK Signaling, mTOR Signaling, and more.
You can also watch an on-demand webinar featuring Dr Reuben Shaw of the Salk Institute for Biological Studies, which focuses on autophagy and ULK1 as a potential therapeutic target.
Select References:
- Shang L, Chen S, Du F, Li S, Zhao L, Wang X. Nutrient starvation elicits an acute autophagic response mediated by Ulk1 dephosphorylation and its subsequent dissociation from AMPK. Proc Natl Acad Sci U S A. 2011;108(12):4788-4793. doi:10.1073/pnas.1100844108
- Yang Z, Klionsky DJ. Eaten alive: a history of macroautophagy. Nat Cell Biol. 2010;12(9):814-822. doi:10.1038/ncb0910-814
- Tsukada M, Ohsumi Y (1993) Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett. 333(1-2), 169–74.
- Kuma A, Komatsu M, Mizushima N. Autophagy-monitoring and autophagy-deficient mice. Autophagy. 2017;13(10):1619-1628. doi:10.1080/15548627.2017.1343770
- Ganley IG, Lam du H, Wang J, Ding X, Chen S, Jiang X (2009) ULK1.ATG13.FIP200 complex mediates mTOR signaling and is essential for autophagy. J. Biol. Chem. 284(18), 12297–305.
- Bach M, Larance M, James DE, Ramm G (2011) The serine/threonine kinase ULK1 is a target of multiple phosphorylation events. Biochem. J. 440(2), 283–91.
- Itakura E, Mizushima N (2010) Characterization of autophagosome formation site by a hierarchical analysis of mammalian Atg proteins. Autophagy 6(6), 764–76.
- Kim J, Kundu M, Viollet B, Guan KL (2011) AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 13(2), 132–41.
- Egan DF, Shackelford DB, Mihaylova MM, Gelino S, Kohnz RA, Mair W, Vasquez DS, Joshi A, Gwinn DM, Taylor R, Asara JM, Fitzpatrick J, Dillin A, Viollet B, Kundu M, Hansen M, Shaw RJ (2011) Phosphorylation of ULK1 (hATG1) by AMP-activated protein kinase connects energy sensing to mitophagy. Science. 331(6016), 456–61.
- Park JM, Lee DH, Kim DH. Redefining the role of AMPK in autophagy and the energy stress response. Nat Commun. 2023;14(1):2994. Published 2023 May 24. doi:10.1038/s41467-023-38401-z
-
Egan DF, Chun MG, Vamos M, Zou H, Rong J, Miller CJ, Lou HJ, Raveendra-Panickar D, Yang CC, Sheffler DJ, Teriete P, Asara JM, Turk BE, Cosford ND, Shaw RJ (2015) Small Molecule Inhibition of the Autophagy Kinase ULK1 and Identification of ULK1 Substrates. Mol. Cell 59(2), 285–97.
- Russell RC, Tian Y, Yuan H, Park HW, Chang YY, Kim J, Kim H, Neufeld TP, Dillin A, Guan KL (2013) ULK1 induces autophagy by phosphorylating Beclin-1 and activating VPS34 lipid kinase. Nat. Cell Biol. 15(7), 741–50.
A version of this blog post originally appeared in September 2017. An updated version was published in May 2024. 24-BRE-71351