We frequently get this question from scientists: “Which species do your PTMScan antibodies work in?”
In antibody‑based assays, species reactivity is a fundamental consideration, whether you’re matching a primary with the appropriate secondary or determining if an antibody against a human protein will recognize the homologous epitope in mouse or another organism. In the context of post-translational modifications (PTMs), there’s an added twist—the amino acid sequence around a PTM site can be conserved, while the modified residue itself is absent or not detectably modified in a given species. For conventional, sequence‑specific antibodies, these constraints determine whether a reagent is usable in your model system.
For CST® PTMScan Kits and PTM antibodies, however, the story is different: these reagents are designed to recognize the PTM itself in a way that is species-agnostic, and can, in principle, enrich modified peptides in everything from model organisms to less commonly studied but experimentally relevant species.
< Jump to the product list at the end of this post >
How PTMScan Antibodies Enable Cross-Species PTM Enrichment
CST proteomics products recognize a specific PTM independently of the surrounding amino acids. The product line includes PTMScan Kits for LC‑MS–based PTM enrichment, as well as antibody reagents and products for western blot–based screening and optimization, both of which rely on the same principle of PTM antibody recognition in different assay formats.
Explore how PTMScan products enable advanced proteomics in the blog, A Revolution in Proteomics Analysis: PTMScan 20 Years Later
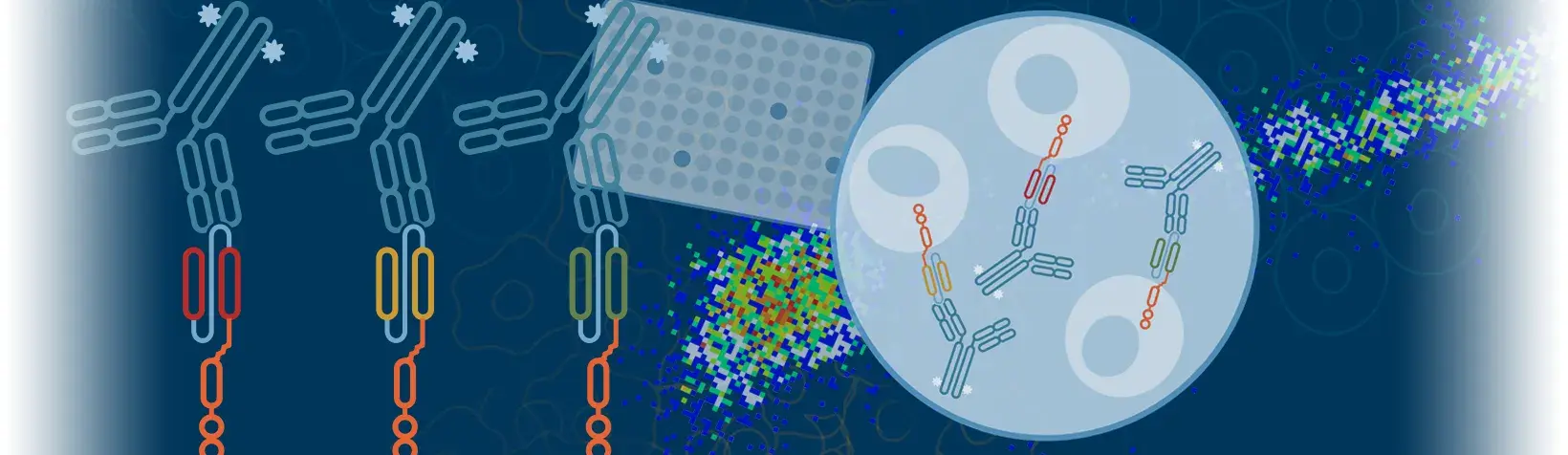
For example, when enriching for tyrosine phosphorylation (pY), the PTMScan Kits contain antibodies that recognize and bind to the phosphate group on a tyrosine residue, regardless of the protein's surrounding amino acid sequence, throughout the proteome (Figure 1, left). Stated differently, CST proteomics antibodies recognize the population of substrates, rather than a specific substrate, that contains the PTM of interest.


This is different from how many other antibodies to PTMs work—most traditional PTM reagents bind to both the modification and the surrounding amino acid sequence, making it specific to a particular post-translationally modified protein or isoform, and therefore likely validated only for a specific species (Figure 1, right).
Depending on your experimental question, there are instances where each type of antibody is useful and relevant for a particular research effort. For example:
- Site-specific PTM antibodies are typically species-specific and can be used to probe a defined PTM site on a particular protein, or a PTM site on a series of protein isoforms.
- CST PTMScan Kits and PTM antibodies recognize only the PTM, and therefore, can be used in any species that expresses sufficient levels of the PTM of interest, as demonstrated in the many research papers highlighted below.
PTMScan Kits are a subset of the CST proteomics portfolio, and they rely on antibodies to enrich PTM peptides for LC-MS–based detection. Since the PTMScan workflow involves denaturing proteins in a urea buffer, disruption of disulfide bonds, and protease digestion, the peptide substrates for the immunoaffinity purification (IAP) largely lack secondary structures that may otherwise influence antibody binding. In theory, this context‑independent recognition means PTMScan antibodies can enrich PTM‑modified peptides from many different species.
In practice, CST scientists test and validate PTMScan Kits using both mouse and human samples during development. Beyond these tested species, however, researchers have demonstrated the universality of PTMScan Kits and proteomics antibodies in a wide variety of organisms.
Published Examples of CST Proteomics Antibodies & PTMScan Kits in Diverse Species
Below is a table listing a selection of recent papers that applied CST proteomics products to study various species besides humans. These studies span vertebrate models, plants, fungi, protozoa, archaea, and bacteria, and collectively demonstrate that these reagents can support cross‑species PTM enrichment when appropriate experimental controls are in place.
Considerations for Using PTMScan Products in New Species
Before embarking on a PTMScan-based LC-MS study of any species, there are a few considerations to take into account.
First, not every species may possess the PTM of interest. For instance, the ubiquitin family that is highly conserved across eukaryotes and yields a diglycine remnant upon trypsin digestion is not present in prokaryotes. Western blot analysis using antibodies against an individual PTM can first confirm that the modification is present in your species or model system. For example, Ubiquitin (E6K4Y) Rabbit Monoclonal Antibody #20326 can be used to assess global ubiquitination levels in any eukaryotic species (Figure 2).

Figure 2. Western blot analysis of extracts from HeLa, NIH/3T3, and C6 cells, untreated (-) or treated with MG-132 (+), using Ubiquitin (E6K4Y) Rabbit Monoclonal Antibody #20326 (upper) or GAPDH (D16H11) Rabbit Monoclonal Antibody #5174.
The same PTM‑directed antibodies can be used for additional analysis to determine how that PTM is modulated across cell lines, treatments, and time points before you commit to a more expensive PTMScan LC‑MS experiment. By probing for changes in signal across conditions, you can cost-effectively identify responsive cell types, select sensitive versus resistant lines, optimize dose and time points for maximum PTM modulation, and determine whether multiple PTMs participate in a stress response or disease process.

For example, Phospho-Tyrosine (P-Tyr-1000) MultiMab® Rabbit Monoclonal Antibody mix #8954 and Acetylated-Lysine Antibody #9441 can be used to assess phosphotyrosine (Figure 3) or acetyl‑lysine (Figure 4) via western blotting, respectively.


Second, processing samples from different species may involve unique technical challenges, such as disruption of cell walls in budding yeast or excess abundance of yolk proteins in Drosophila embryos. No matter the biochemical workflow optimized for the particular organism, it is important to preserve PTMs through the use of the appropriate inhibitors (for example, Phosphatase Inhibitor Cocktail #5870) and to minimize artifactual modifications during processing that complicate LC-MS analysis, such as lysine carbamylation arising from heating samples in urea buffer. The use of denaturing buffer conditions, such as 8M urea, can facilitate the homogenization and extraction efficiency of challenging specimens.
Additionally, the use of spike‑in positive control peptides containing the PTM of interest can help validate the IAP procedure when initiating work on an unfamiliar species and increase confidence in cross‑species comparisons.
Blog: Reproducibility in Proteomics Experiments: Using Control Peptides with PTMScan
Finally, having a well-annotated database of all protein sequences from that species is essential for the bioinformatic analysis of the LC-MS data. A poorly annotated or incomplete database will hamper the identification of potential PTM sites, although advancements in de novo mass spectrometry searches that do not rely on a protein database may help in these contexts.
PTMScan Kits & PTM Antibodies Are Validated for Cross‑Species PTM Profiling
In short, CST PTMScan Kits and proteomics reagents have established compatibility with a wide range of species, illustrating the versatility of the reagents as a general tool to leverage PTM proteomics for monitoring cellular signaling and protein regulation. The combination of PTM antibodies, denaturing IAP workflows, and LC‑MS analysis makes PTMScan Kits and proteomics antibodies well‑suited for PTM profiling across many species.
For researchers working in less common species or emerging model organisms, these data support the use of these powerful reagents as a practical starting point for cross‑species PTM enrichment.
Proteomics Antibodies & PTMScan LC-MS Kits from CST
The table below includes a list of select CST proteomics solutions for the species-agnostic detection or enrichment of the common PTMs: acetylation, phosphorylation, and ubiquitination.
There are dozens more proteomics solutions in our catalog for additional PTMs—reach out to us for help navigating our extensive portfolio!
| PTMScan Kits for LC-MS Analysis | PTM Antibodies for Western Blotting |
| Acetylation | |
|
|
| Phosphorylation | |
| Ubiquitination | |
|
Will require the use of a species-specific or cross-reactive antibody for ubiquitin. |
|
|
Dozens more PTMScan Kits and PTM antibodies for additional PTMs are available in the CST product catalog — and don't hesitate to reach out to us for one-on-one support! |
|
26-TPD-47850